文章題目:A plasma membrane transporter coordinates phosphate reallocation and grain filling in cereals
磷(Pi) 對植物生長和作物產量至關重要。然而,在籽粒灌漿過程中 Pi 穩態是如何維持的還不清楚。在本研究中,何祖華課題組通過圖位克隆鑒定了一類關于水稻籽粒灌漿調控 Pho1 -Pi 的品系:OsPHO1;2。且發現該品系在灌漿過程中 Pi 重新分配中的特殊作用有關。 伯豪生物為何祖華課題組提供了 RNA-seq 服務,發現 PHO1;2 通過影響 AGPase 的活性進而影響水稻灌漿過程中 Pi 重新分配的作用機制。AGPase 對淀粉合成至關重要,而在 Ospho1;2 突變體植株中,AGPase 的過表達緩解了籽粒灌漿缺陷。該機制在玉米中得到了證實, 為在谷物生長過程中以較小的磷肥投入來提高籽粒產量,提供了有效方案。
籽粒灌漿突變體 OsPHO1;2 的表型及圖位克隆
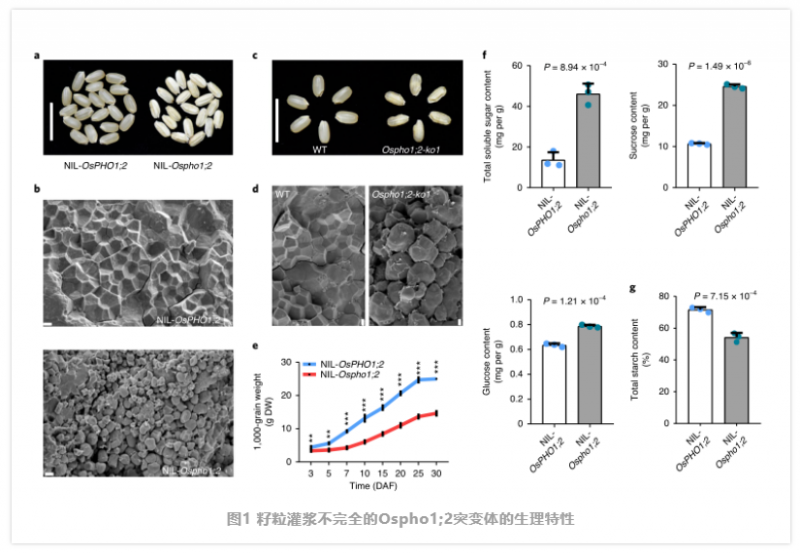
何祖華課題組發現了一個自然發生的籽粒灌漿較差表型的水稻突變體品系。該缺陷表型由一個隱性位點控制,經過基因克隆后分別命名為 NIL-OsPHO1 和 NIL-OsPHO2。NIL-OsPHO2 的籽粒表現出典型的灌漿缺陷,淀粉粒松散、排列不整齊,導致籽粒變薄、皺縮,造成嚴重的產量損失。除分蘗數減少(圖 1) 外,其余突變體植株正常。在整個種子發育過程中,植株的千粒重減少了近一半(圖 1e)。與 gif1 突變株不同,NIL-OsPHO1 的淀粉含量更少,可溶性糖更多,對白葉枯病的抗性更強(圖 1)。這表明,這兩種突變體具有不同的生理表型。在該區域作者鑒定了一個單基因 LOC_Os02g56510,該候選基因編碼 PHO1 家族的 Pi 轉運蛋白。為了確定 NIL-OsPHO1;2 的原因突變,作者對其 5kb 區域進行了測序,發現在 OsPHO1;2 的第 7 外顯子中有 1 -bp 的缺失,導致編碼序列的幀移和過早終止。作者還在第 1 外顯子和第 3 外顯子中發現了兩個同義 SNP,在第 12 內含子中發現了一個 SNP。為了確定這一缺陷灌漿表型是否是由 OsPHO1;2 突變引起的,作者在 Nipponbare (NIP) 中構建了 8 個 CRISPR/Cas9 敲除株系,分別為 OsPHO1;2 -ko1 到 OsPHO1;2 -ko8,這些株系均表現出與 NIL-Ospho1;2 相同的灌漿缺陷(圖 1)。
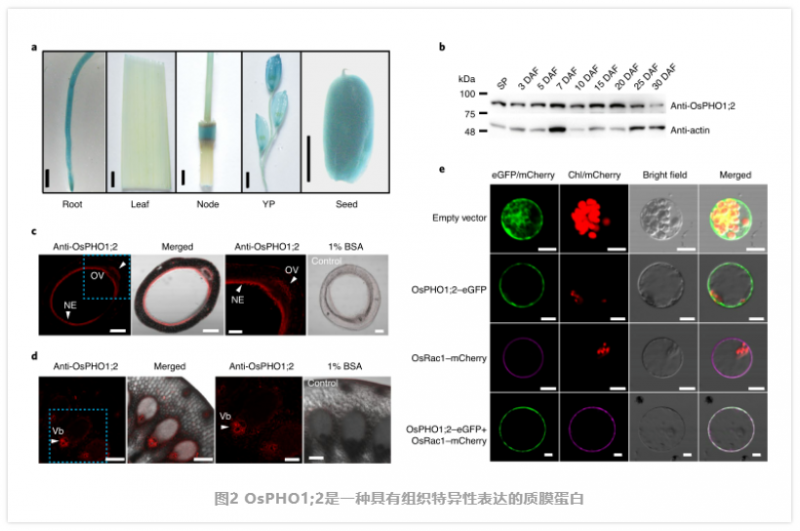
PHO1;2 是一種在種子發育過程中高度表達的質膜蛋白。通過定量反轉錄 PCR (qRT - PCR) 發現,除了在幼穗、外殼和根中表達外,OsPHO1;2 在發育中的種子中也有高表達。通過對轉基因植物中 2 -β- 葡萄糖醛酸酶(GUS) 報告基因的組織學分析,證實了該表達模式(圖 2a)。同樣,從開花后 0 d(穗)到 30 d( 穗)的整個灌漿階段,OsPHO1;2 蛋白均在籽粒中富集。為了進一步研究 OsPHO1;2 的細胞特異性,作者在籽粒灌漿早期的種子樣本中進行了抗 OsPHO1;2 抗體的免疫熒光試驗。有趣的是,在種子的珠心表皮(NE) 和胚珠脈管系統(OV) 中檢測到強烈的熒光信號,這些細胞起著向胚乳裝載和排出營養物質的作用 m3(圖 2c)。在 I 節維管束中也發現了熒光信號(圖 2d),這有助于水稻植株中的養分分布。作者進一步驗證了 OsPHO1;2 在發育中的種子不同組織中的表達,發現在胚胎、胚乳、果皮和 NE 中都可以檢測到 OsPHO1;2 mRNA 和 OsPHO1;2 蛋白。此外,利用含有 pOsPHO1;2- Gus 報告基因的轉基因株系進一步驗證了 PHO1;2 在種子中的定位。作者將一個瞬時表達的綠色熒光蛋白(GFP) 與 OsPHO1;2 融合進入水稻鞘原生質體和洋蔥表皮細胞,發現在這兩種情況下,OsPHO1;2 - GFP 主要定位于質膜(圖 2)。細胞內偶爾可見微弱的熒光信號,提示部分 OsPHO1;2 蛋白可能定位于細胞內隔間。綜上所述,這些結果表明,OsPHO1;2 是一種質膜定位蛋白,在發育中的種子 NE 和維管組織中高表達。
OsPHO1;2 是 Pi 的轉運體
由于沒有直接證據表明植物 Pho1 型蛋白,如 OsPHO1;2 等是 Pi 的轉運體。值得注意的是,雖然 Ospho1;2 幼苗比野生型(WT) 幼苗生長早期,可能由于缺陷胚乳提供的能量較少,它們后來恢復了 WT 的高度。這些生長動態表明,OsPHO1;2 可能不是整個植物生長發育所必需的,但在籽粒灌漿過程中是必不可少的。為了分析 OsPHO1;2 蛋白在籽粒灌漿和 Pi 穩態中的作用機制,作者試圖通過三種不同的異源表達系統來確定 OsPHO1;2 蛋白是否確實作為 Pi 轉運體。首先,作者發現 OsPHO1;2 補充了酵母突變體 EY917 的生長表型,該突變體缺乏 5 個負責 Pi 攝取的轉運體(pho84?,pho87?,pho89?,pho90?,pho91?),這證明了 OsPHO1;2 可能具有 Pi 轉運活性。主要的外排活性可能介導了木質部的 Pi 負荷,從而解釋了先前的遺傳分析顯示它們在根到地上部 Pi 轉移中的作用。為了將 OsPHO1;2 在 Pi 轉運中的作用與 Pi 穩態聯系起來,作者比較了近距離接觸的幼苗之間的 Pi 水平。在缺 Pi 或正常供應 Pi 條件下,突變株根系中 Pi 含量增加,而地上部 Pi 含量降低。因此,OsPHO1;2 突變損害了 Pi 從根到莖的轉移,這與之前的結果一致。此外,31P NMR 分析顯示,突變體幼苗根細胞中胞質和液泡中 Pi 含量明顯較高,排除了 OsPHO1;2 參與胞質 - 液泡劃分的可能性(圖 3)。接下來,作者研究了成熟期植株地上部組織中 Pi 的分布,發現在節、殼和糙米中,NIL-Ospho1;2 和 Ospho1;2-ko1 植株中 Pi 的含量均高于 WT 植株,但葉片中 Pi 的含量較低。表明 OsPHO1;2 可能參與了 Pi 在種子和葉片之間的重新分配。
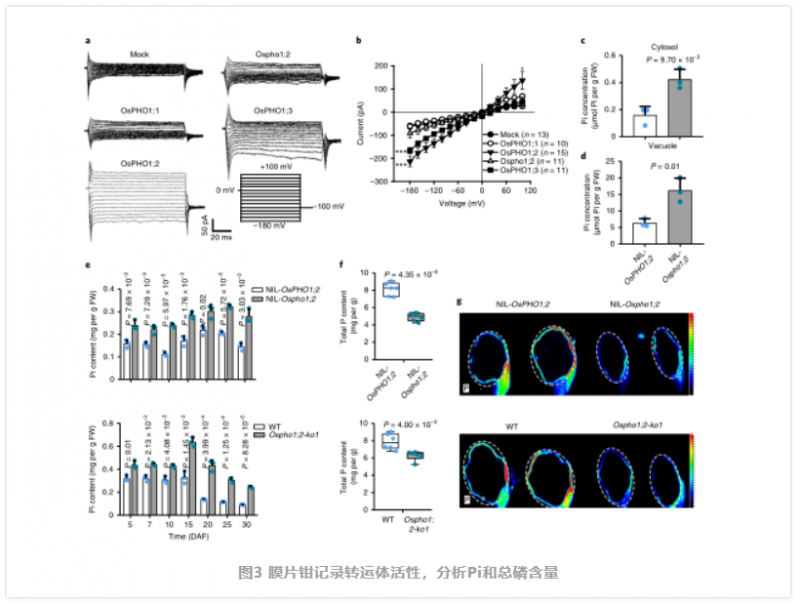
為了支持上述假設,作者進行了測量在整個灌漿過程中發現該突變體的 Pi 含量較高(圖 3e)。由于 OsPHO1;2 在發育中的種子 NE 組織中高度表達,我們比較了 NE 周圍胚乳和果皮中 Pi 的含量,發現在 Nilosho1;2 種子中,這些組織中 Pi 的含量顯著增加。因此,在 Ospho1;2 突變體中,Pi 的再分配受到了干擾。而突變體種子中總磷含量顯著降低(圖 3f),微 x 射線熒光(μXRF) 成像結果一致(圖 3g)。這說明突變株的有機磷合成可能由于種子中母、子組織間 Pi 交換的中斷而減少。因此,我們得出結論,OsPHO1;2 作為 Pi 外排轉運體,將 Pi 從胚乳細胞中釋放出來,其功能的喪失導致 Pi 在種子中過量。過量的 Pi 積累抑制淀粉合成酶。為探討灌漿期 Pi 積累與灌漿之間的關系,作者對灌漿期 NILs 進行了 RNA-seq 比較分析,鑒定出大量差異表達的基因。作者發現許多參與淀粉和糖代謝的基因被富集。此外,作者通過 qRT-PCR 確定了幾個與淀粉合成相關的代表性基因的表達模式,包括編碼 AGPase、淀粉合酶、淀粉分支酶和淀粉去分支酶 29(圖 4a)。這些基因在 Ospho1;2 突變體中大部分被下調,這與該突變體在淀粉合成方面的缺陷相一致。免疫印跡和同工酶分析顯示,突變體種子中參與淀粉合成的幾個關鍵酶的蛋白水平或酶活性也有所下降(圖 4)。在這些酶中,AGPase 催化淀粉合成的步驟一,從 G -1- P 和 ATP 中生成 ADP-Glc 和 PPi, AGPase 的功能障礙會導致籽粒灌漿缺陷,這與 Ospho1;2 突變體中發現的缺陷類似,表明 AGPase 在籽粒灌漿過程中起著不可或缺的作用。
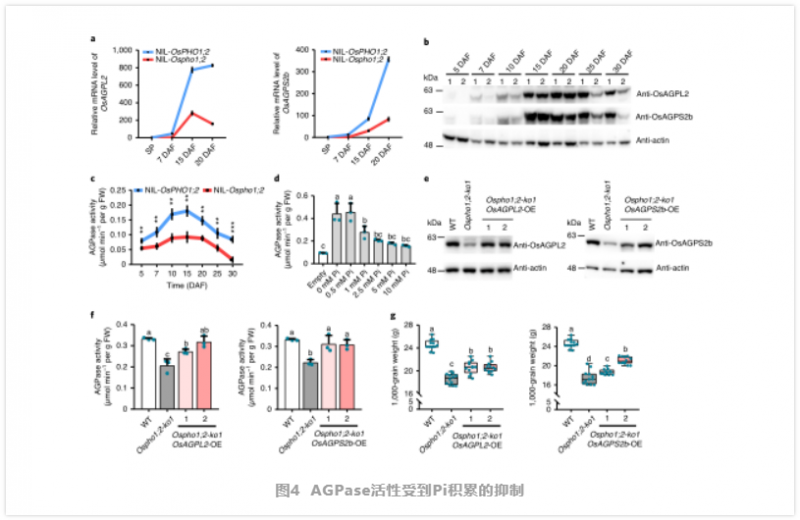
綜上所述,作者發現過量的 Pi 水平對 AGPase 的 mRNA 水平和活性都有負面影響,這可能是 Ospho1;2 突變體淀粉合成減少和籽粒灌漿缺陷的基礎。為了進一步闡明 AGPase 活性降低與籽粒灌漿缺陷之間的因果關系,作者在 Ospho1;2-ko1 背景下的玉米泛素啟動子驅動下,異位過表達了 OsAGPL2 和 OsAGPS2b。選擇兩個過表達每個轉基因的純合子系進行實驗,每個純合子系的蛋白質水平都恢復到相對于 WT 的水平。作者發現,AGPase 過表達(OE) Ospho1;2-ko1 系的 AGPase 活性低于 WT,但高于 Ospho1;2-ko1 突變體的 AGPase 活性(圖 4)。在收獲后,我們觀察到 AGPase-OE Ospho1;2-ko1 系的灌漿缺陷得到了部分補救,而過表達 OsAGPL2 或 OsAGPS2b 可以顯著增加千粒重(圖 4)。因此,這些結果進一步支持了 ZmPHO1;2 在籽粒灌漿和 Pi 重新分配中起著重要作用。
伯豪生物服務產品助力國內外臨床科研單位發表 SCI 論文 1900 余篇,雜志總影響因子超過 10000 分,其中 Nature、Science、Cell、Lancet Oncol 等高水平論文等高水平論文超過 50 篇。影響因子大于 5 分的文章 659 篇,影響因子大于 10 分的文章 255 篇,影響因子大于 20 分的文章 78 篇,文章領域涉及腫瘤、免疫、神經、發育等多個領域。

更多伯豪生物人工服務:

